
[ad_1]
In the great diversity of microbial life, there are many examples of symbiosis – a process by which different organisms maintain a stable relationship with each other. Such interactions generally provide some form of benefit to one or more of the organisms involved. Write in Nature, Graf et al.1 report the discovery of a fascinating example of symbiosis between microorganisms. This finding could shed light on the types of processes that led to the evolution of mitochondria, the energy-producing organelles found in eukaryotic cells (those containing a nucleus).
In some cases, the symbiotic partners can integrate in such a way that one partner is taken up into the cell of the other partner through a process called endosymbiosis (Fig. 1). Endosymbiotic interactions have enabled key transitions in the history of life on Earth. The interaction that gave birth to the first eukaryotic cell is believed to be a prime example. This ancestral eukaryote probably formed from an endosymbiosis in which a host cell belonging to a group of unicellular organisms called Asgard archaea2,3 took on a type of bacterial cell belonging to the phylum Proteobacteria4. The mitochondria of eukaryotic cells are believed to be the direct descendants of these once free proteobacterial endosymbionts.
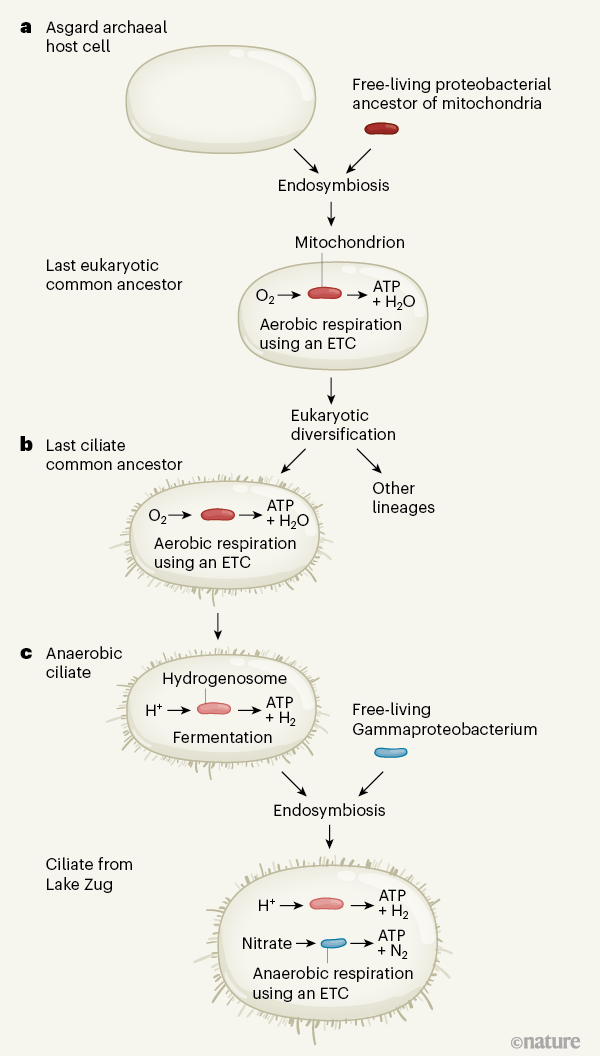
Figure 1 | Evolution of Organelle. a, Eukaryotic cells, those with a nucleus (nucleus not shown), probably arose when a type of single-celled host called an archaial Asgard cell2,3 took a proteobacterial cell in a process called endosymbiosis4. The proteobacterium evolved to form mitochondrial organelles in the last common ancestor of all eukaryotes. Through the process of aerobic respiration, in which oxygen (O2) is consumed and water is formed when electrons are transferred along an electron transport chain (ETC), a mitochondria produces energy in the form of ATP molecules. bEukaryotes have diversified and only a few lineages, such as the last common ancestor of organisms called ciliates, have retained mitochondria. vsSome ciliate lines adapted to thrive in oxygen-free environments. The mitochondria of these ciliates evolved into organelles called hydrogenosomes. Anaerobic ciliates perform fermentation, a process in which ATP is generated and hydrogen ions (H+) gain electrons to form hydrogen (H2). Graf et al.1 report the discovery of an anaerobic ciliate with a gammaproteobacterial endosymbiont. The genome of this bacterium encodes the components necessary to generate ATP by converting nitrate into nitrogen (N2) using anaerobic respiration and to export ATP to the host. This discovery reveals how anaerobic ciliates can regain the ability to practice breathing using an ETC. These characteristics are similar to the processes associated with mitochondrial evolution.
Graf and his colleagues describe an intriguing bacterial endosymbiont they discovered living in a ciliate, a type of single-celled eukaryotic microbe. Although associations between these types of organisms are not uncommon, this particular case exhibits some of the hallmarks of the type of endosymbiosis that gave rise to mitochondria. This includes the production of energy in the form of ATP molecules through the process of energy generation known as respiration. This process is also connected to a mechanism by which ATP could be exported from the endosymbiont to provide energy to the host.
The environmental conditions and atmospheric composition on Earth today differ considerably from those that existed when life first appeared on this planet more than four billion years ago. During what is known as the great oxidation event, about 2.1 billion years ago5, oxygen accumulated in the atmosphere and ocean waters, a development that has had a major impact on the course of life on Earth. Oxygen is toxic to most organisms that thrive in environments lacking it. However, some microorganisms have learned to harness the chemical properties of oxygen, using it as an electron acceptor in energy production pathways. This type of oxygen-dependent process, called aerobic respiration, is much more energy efficient than fermentation, an ancient form of non-oxygen-dependent energy metabolism that allows many anaerobic organisms to survive without oxygen.
Eukaryotic cells probably appeared some time after the great oxidation event6, and their ability to perform aerobic respiration was acquired by the mitochondrial endosymbiont. It is believed that the efficient energy production capacity this provided conferred selective advantages during eukaryotic evolution, although the exact contribution, for example, to the emergence of cellular complexity is discussed.7.
Current evidence6 indicates that the evolution and diversification of early eukaryotes occurred under conditions in which oxygen was present. However, some groups of eukaryotes nonetheless thrive in oxygen-deprived environments. It is believed that these anaerobic eukaryotes evolved from aerobic ancestors carrying mitochondria.
In the absence of oxygen, anaerobic eukaryotes use a fermentation-based metabolism, which subjects them to more stringent energy regimes than those of aerobic eukaryotes. This type of metabolism has appeared in various types of anaerobic eukaryotes and is associated with the evolution of mitochondria into organelles called hydrogenosomes.8. All hydrogenosomes have to some extent lost the mitochondrial pathways for aerobic respiration and generate hydrogen rather than carbon dioxide and water as the end product of their energy generation pathways. Ciliates are exceptionally efficient at adapting to oxygen-depleted environments, and mitochondria-hydrogenosome transitions have occurred independently several times in this group of organisms.9.
Graf and his colleagues studied an anaerobic ciliate belonging to the class Plagiopylea, found in the deepest layers of Lake Zug in Switzerland. This environment lacks oxygen and contains relatively high levels of nitrate. An initial microscopic evaluation revealed that, unusually, these ciliates have a bacterial endosymbiont (belonging to the Gamma-Proteobacteria class), rather than an archaic endosymbiont which produces methane, which is the most typical type of endosymbiont. present in anaerobic ciliates.
DNA sequencing data for the lake water samples revealed the presence of genes indicating that hair cells have hydrogenosomes. Additionally, sequencing data indicates that bacterial endosymbionts have an electron transport chain – a collection of protein complexes for respiration that are used to generate energy through a process called oxidative phosphorylation. Graf et al. propose that the electron acceptor in this chain is nitrate, rather than oxygen used by aerobic organisms. Consistent with this model, the authors report that denitrification rates (the microbial process that converts nitrate to nitrogen) were higher in lake water samples in which ciliates were present than in those with ciliates removed. .
The endosymbiont genome identified by Graf and colleagues is significantly smaller than the genomes of most microbial eukaryotic endosymbionts, containing just 310 protein-coding genes. Among these, the authors identified a gene that encodes a potential transport protein for ATP, which they suggest is used to export ATP from the endosymbiont to its host, allowing the ciliate to utilize the endosymbiont for energy production by “ breathing ” nitrate. This finding represents a unique example of an endosymbiont that contributed to the respiration capacity (albeit using nitrate instead of oxygen as an electron acceptor) in a eukaryote that apparently retains organelles of mitochondrial origin ( hydrogenosomes) – ancestral versions of which once performed respiratory functions. functions.
Curiously, several parallels can be drawn between the cellular partnership discovered by Graf and his colleagues and the evolution of mitochondria in eukaryotes. In both cases, the respiratory capacity was acquired by an anaerobic host cell through the metabolic integration of a proteobacterial endosymbiont, and mechanisms can be identified for the energy exchange between the symbiont and the host cell. In addition, a substantial reduction in the endosymbiont genome is observed, although mitochondrial genomes are generally either much smaller than the endosymbiont genome observed by Graf and colleagues, or have been completely lost (as is the case for several hydrogenosomesten).
Despite these fascinating similarities, there are also some notable differences. Mitochondrial endosymbiosis was a much older event, involving an archaeal host cell rather than a modern eukaryotic cell. Mitochondria, although now reduced from their original form, or even lost from some current eukaryotes11, has become an integral part of eukaryotic cells. Genes inherited from the original mitochondrial endosymbiont were often re-targeted to the nuclear genome, and some of the proteins encoded by these genes took on various functions throughout the cell. A similar level of integration is unlikely for the bacterial endosymbionts of the ciliate studied.
Nonetheless, it would be interesting to study whether there has been any relocation or reuse of genes between host and endosymbiont, and to what extent a typical mitochondrial function, such as ATP production, has been overridden. or preserved in the hydrogenosomes of the ciliate. Evidence indicating that the ATP transporter identified by Graf and his colleagues can export ATP to the ciliated host would help confirm the proposed symbiotic interaction. The further discovery and exploration of equally surprising symbiotic interactions in poorly explored parts of the microbial world is certainly an exciting prospect for the future.
[ad_2]
Source link